A PSA szintek lognormális eloszlása: a PSA releváns paramétereinek értékelése
Elvileg két legegyszerűbb lehetőség: ezek a paraméterek
- bevezetni (szabálytalan) zajt, amely elrejti a PSA függését a ráktól vagy
- a PSA-koncentráció szisztematikus megváltoztatása, amely elválasztható a rák hatásától.
Punglia et al. a BPH hatásának a PSA-koncentrációra gyakorolt hatásának értelmezésére használt második megközelítést alkalmazta. Ebben a tanulmányban javaslom a prosztata stimuláció hatását ugyanúgy értékelni.
Bevezetés:
A lognormális PSA koncentráció eloszlása a különféle BPH-ról a rákról
Valószínűségi eloszlás és vevő funkciója
f a megfelelő Valószínűségi eloszlás. Definíció szerint az f alatti terület (az “infinity” és “infinity” között) 1.
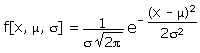
A Vevőegység jellemzője (ROC) egy parametrikus görbe, melynek paramétere a teljes PSA koncentráció psa (“küszöb”). A ROC görbe pontjai a koordinátákkal rendelkeznek , ahol a TP és az FP a normál eloszlású f [x, 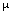 ,
, 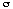 ] a küszöbérték jobb oldalán x = ln [psa].
] a küszöbérték jobb oldalán x = ln [psa].
- TP [psa] a megfelelő terület az f alattk, a rákos populáció eloszlása. (“valódi pozitív frakció”).
- FP [psa] a megfelelő terület az f alattg, a rák nélküli népesség eloszlása (“hamis pozitív frakció”).
A ROC kifejezése a normál eloszlások szempontjából
Mivel , és így a ROC, az elsődleges információ az orvosi adatokból (azaz a PSA mérésekből és a biopsziákból), a lognormálisg és fk a numerikus illeszkedés eredménye. Ábrákon. Az 1. és a 3. ábra megmutatja, hogy ez hogyan történik, és az eredményt, az eloszlást fg és fk, az ábrák bal alsó sarkában látható árnyékolt négyszögben jelennek meg.
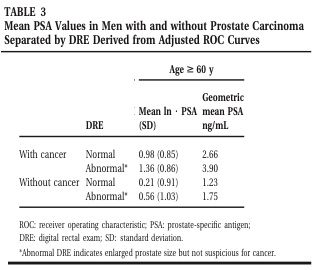
A 3. táblázatban Punglia et al. adja meg a kapott terjesztések paramétereit ( = átlagos ln [PSA], = szórás). A k és g populációkat tovább osztották két szubpopulációra a digitális rektális vizsga (DRE) eredményei alapján: DRE normál és DRE abnormális.
1.ábra. ROC görbék csatlakoztatása (jobb felső sarok) lognormálisan elosztott PSA koncentrációkkal (árnyékolt terület a bal alsó sarokban). Az árnyékos területen eloszlások az alpopulációhoz
- a rendes (rendellenes) DRE-t a telek felső (alsó) felében mutatják,
- rák nélkül (rák) a g (k) szilárd (szaggatott) görbék képviselik.
Az 1. ábra mutatja: A BPH jelenlétét
- kevés hatással van a szilárd és szaggatott görbék alakjára és relatív pozícióikra, így a ROC-görbére,
- a szilárdtestű görbeegyüttest a magasabb PSA szintek felé tolja át, ezáltal megváltoztatva a PSA küszöbcímkét a ROC görbén.
Ebben az értelemben a BPH hatása a PSA-koncentrációra a rák hatása fölött helyezkedik el, ahelyett, hogy elfedné a hatását. (Ezzel szemben a nukleáris mágneses rezonancia leképezésben (MRI) a BPH és / vagy a krónikus prosztatitis rákot maszkolnak [Shukla-Dave A és munkatársai, “Krónikus Prostatitis: MR Imaging és 1 H MR Spektroszkópos Imaging Observations – Initial Observations” , Radiology 2004. június 231 (3): 717-24 (gyorsítótárban)]
A lognormális PSA koncentráció eloszlása a rákos mechanikus prosztata irritációval szemben
A szexuális aktivitás szintjét megfigyelték, hogy korrelál a tPSA-koncentrációkkal. A megfelelő tPSA magasság körülbelül azonos méretű lehet, mint Punglia et al. mutatott rá a rákra. A szexuális aktivitás csökkentése nem fogja könnyen csökkenteni a tPSA szinteket. A tPSA relaxáció hetekig tarthat. Ez meghaladja az urológusok ajánlását, hogy pácienseik PSA mérés előtt több napig szexuálisan állóképesek legyenek.
Itt javasoljuk, hogy a szexuális aktivitás PSA-tesztre gyakorolt hatását hasonlóan kezeljük, mint a BPH egyikét, vagyis a g és k lognormális eloszlások eltolódását. A terv utolsó részében két lehetőség fog tárgyalni:
- Ha a szexuális aktivitás hasonlóan hat a BPH-hoz, a PSA-tesztre gyakorolt hatása csökkenthető a tesztküszöb beállításával.
- Ha a szexuális aktivitás változatlanul hagyja a PSA-koncentráció valószínűségi eloszlásának helyét a rák esetében, a PSA-vizsgálat nem különböztethet meg szexuális aktivitást és rákot.
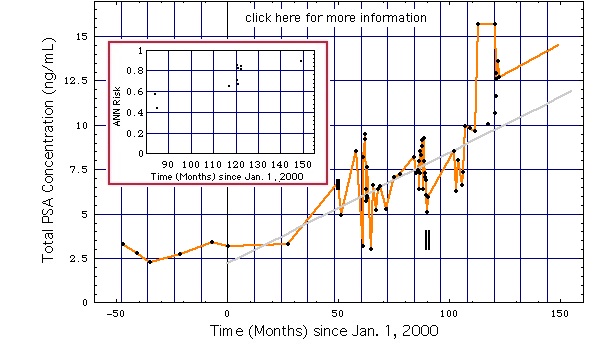
bal diagram: A PSA-koncentráció rezgése valószínűleg a szexuális aktivitás változásai (adatok) miatt következett be.
- Az adatpontok egyenes vonalakat önkényesen összekapcsolják.
- Függőleges rudak:
- az 50-es vagy 75-ös havi 50. vagy 75. havi görbén: hosszúság = 2 x 0,2 ng / ml = 2 x standard szórás, a PSA-koncentráció mérési együtthatójának (kevesebb mint 3% -a) alapján számítva a 28-as munkacsoport Angaben zur Messunsicherheit.
- A 90-es hónap múlva egymás melletti 2 vonal jelöli azokat az időpontokat, amikor a Mágneses Resonancia Imaging (MRI) és a Fine Needle Aspiration Biopsy (FNAB)
- MRI és MRSI bal oldali vonallal jelölt időpontban. MRI eredmény: nincs prosztatarák.
- FNAB (a DNS-ploiditás analízissel), amelyet a jobb oldali vonallal jelöltek. Biopsziás eredmény: nincs prosztatarák.
- A szürke vonal 0,75 ng / ml-es dőlésszög alatt a Coakley FV, Chen I, Qayyum A, Westphalen AC, Carroll PR, Hricak H, Chen MH, Kurhanewicz J. után a küszöbérték PSA növekedését jelzi a prosztata-specifikus antigén érvényessége tumor marker a prosztatarákban szenvedő férfiaknál éber-várakozással: korreláció a sorozatos endorectalis mágneses rezonancia képalkotással és spektroszkópos képalkotással. BJU Int. 2007 Jan; 99 (1): 41-5 (több).
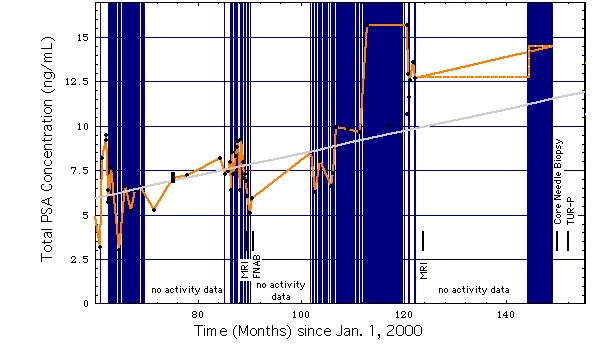
jobb diagram: Ugyanaz, mint a bal oldali diagram, kivéve a magasabb időbeli felbontást és a szexuális aktivitás gyakoriságának grafikus ábrázolását vagy a kiterjesztett kerékpározást. A függőleges (kék) vonalak a szexuális aktivitással vagy a kibővített kerékpározással jelölt napokat jelölnek (azaz nincs biciklizés / szexuális aktivitás jelöletlen napokon). “nincs tevékenységre vonatkozó adat”: ezen időszak alatt a szexuális tevékenységet / kerékpározást nem rögzítették.
Figyelembe véve az átlagos PSA-koncentráció emelkedését a korral, a 2. ábrán a PSA-koncentrációt megközelítőleg 5 ng / ml-es átlagértékkel közelítjük, amely körül nagyjából egy faktor 2, azaz 2,5 ng / ml-től 10 ng / ml-ig. Ezt a szexuális aktivitással rétegzett ROC görbék felépítésére használják.
alternatívák
Ábrákon. 3 a lognormális PSA disztribúciók alakja azok, amelyeket Punglia et al. a normál DRE esetében. A megnövekedett szexuális aktivitást figyelembe veszik a megfelelő eloszlás felfelé történő elmozdításával az említett 2. faktorral. (Megjegyzés: ln2 = 0,7)
1. alternatíva
Alkalmazott paraméterek:
- Az átlagos populáció (rákkal és anélkül) átlagos lnPSA-értékei megfelelnek a megfelelő eszközöknek, amelyeket Punglia et al. a DRE normális populáció számára.
- A “megnövekedett nemi populáció” (rákkal és anélkül) átlagos lnPSA értéke 0,7-nél magasabb, mint a Punglia et al. a DRE normális populáció számára.
Aztán a
- a ROC-görbe alakja – ami a tesztteljesítményt jelenti – független a szexuális aktivitástól (így az átlagos és megnövekedett szexi ROC görbék egybeesnek), és
- A POC küszöbértékek a ROC görbe mentén (szürke számok) nőnek a szexuális aktivitással, példánkban egy tényezővel 2.
Példa: Válasszuk ki a pontot = a ROC-on. A tesztérzékenységet TP = 0,4 és a vizsgálati specifitás (1 – FP) = 0,87
- a vizsgálati küszöbérték x = 3,3 ng / ml az átlagos populációra,
- a vizsgálati küszöbértéknél 2 x = 6,6 ng / ml a megnövekedett nemű alpopuláció esetében.
Eredmény: A megnövekedett szexuális aktivitással rendelkező alpopuláció esetében a PSA-küszöböt a normál populációhoz képest 2-szeresére kell emelni, ami a példánkban azt jelenti, hogy a vizsgálat ugyanazt eredményezi az alpopuláció tagjának, amelynek PSA szintje magasabb 6,6 ng / ml, mint a normál populáció egy tagjának, amelynek PSA szintje 3,3 ng / ml fölött van.
2. alternatíva
Alkalmazott paraméterek:
- Az átlagos populáció (rákkal és anélkül) átlagos lnPSA-értékei megfelelnek a megfelelő eszközöknek, amelyeket Punglia et al. a DRE normális populáció számára.
- A “megnövekedett nemi populáció” átlagos lnPSA értéke
- a rákot úgy választjuk meg, mint a Punglia et al. a DRE normális populáció számára.
- rák nélkül 0,7-nél magasabb értéket kapunk, mint a megfelelő értékek, amelyeket Punglia et al. a DRE normális populációhoz, amint azt a 2. ábrán látható statisztikákból nyerjük ki.
Az alpopuláció ROC gyakorlatilag megegyezik az átlóval (szaggatott vonal a 3b. Ábra ROC diagramjában). A nagy összes PSA-koncentráció (> 10 ng / ml) alatti populációk általában olyan g és k eloszlásokkal rendelkeznek, mint ezek, a szexuális alpopulációhoz képest, mivel PSA-tesztjeik alacsony teljesítményt mutatnak, specifikus érzékenységi tartományukkal (fekete vízszintes vonalak a érzékenység 0,9 és 0,95 a lognormális ábrán b) érintse meg az átlót. Csak akkor, ha több Pca specifikus paramétert is figyelembe veszünk, mint például a mesterséges neuronhálózatban vagy a nomogramban, a tesztteljesítmény növekedése (színes vízszintes vonalak 0,9 és 0,95 érzékenység esetén), mert az egészséges és a beteg mozgása eloszlik.
Eredmény: A két alternatíva esetében a PSA tesztnek nincs megkülönböztető ereje az alpopulációnak a megnövekedett szexuális aktivitással: A PSA tesztben a szexuális aktivitás növekedése nagyon hasonlít a rákra.
Tentatív modell
A PSA összes kémiai formájának koncentrációja a prosztatában sok nagyságrenddel magasabb, mint a vérben. Tegyük fel, hogy
- a vér-prosztata-gát fenntartja ezt a koncentrációs lépést, és
- a vér emelkedett tPSA-koncentrációja a vér-prosztata gáton,
akkor a prosztatarák és az emelkedett szexuális aktivitás egyaránt csökkenti a vér-prosztata gátat.
Vér-prosztata-gát

A gasztrointesztinális gát
A gasztrointesztinális nyálkahártya akadályt képez a test és a lumenális környezet között, amely nemcsak tápanyagokat tartalmaz, hanem potenciálisan ellenséges mikroorganizmusokkal és toxinokkal terhelt. A kihívás az, hogy lehetővé tegye a tápanyagok hatékony transzportját a hámban, miközben szigorúan kizárja a káros molekulák és organizmusok állatba való átjutását. A gyomor és a bél nyálkahártyájának kizáró tulajdonságait “gyomor-bélrendszeri gátként” nevezik.
Nyilvánvaló, hogy számos primer gyomor-bélrendszeri betegség a nyálkahártya-gát megzavarásához vezet, lehetővé téve a szisztémás betegség fokozódását. Ugyanilyen egyértelmű, hogy sok szisztémás betegség folyamat a gasztrointesztinális gát károsodásához vezet, ezáltal további sérelmet okozva egy már kompromittált rendszerben. A gát természetének megértése segíthet az ilyen események előrejelzésében és a profilaktikus vagy aktív terápiák támogatásában.
A gasztrointesztinális gáton gyakran tárgyalták, hogy két összetevőjük van:
A belsõ akadály az emésztőcsövet bélelő hámsejtekből és a szoros összekötő csomópontokból áll.
A belső gyomor-bél rendszeri akadály
Az étkezési csatornát hámsejtek bélelik, amelyek a nyálkahártya meghatározó struktúráját alkotják. Néhány kivétellel a hámsejtek a gyomorban és a belekben körkörösen egymáshoz kötődnek, szoros csomópontokkal, amelyek lezárják a paracelluláris tereket, és ezáltal létrehozzák az alapvető gasztrointesztinális gátat. Az emésztőrendszerben az érintetlen hám fenntartása kritikus fontosságú a gát integritása szempontjából. Általában véve a toxinok és a mikroorganizmusok, amelyek képesek megsérteni az egyetlen réteg epiteliális sejteket, akadálytalanul hozzáférnek a szisztémás keringéshez.
Amint várható, a különböző típusú hámsejtek között sokféle diverzitást mutatnak specifikus gátló funkciókban. Például a gyomor-parietális és fősejtek apikális plazmamembránjai a protonoknál alacsony a permeabilitást mutatnak, ami elősegíti a savak hátsó diffúziójának a sejtekben való károsodását. A kis intesztinális hámsejtekből hiányzik ez a speciális képesség, és így sokkal hajlamosabbak a savval indukált károsodásra.
A gasztrointesztinális hámsejteket körülvevő szoros csomópontok a belső gát kritikus elemei. Ezeket a struktúrákat a hegesztésekhez hasonló passzív szerkezeteknek tekintették, ám a közelmúltban végzett vizsgálatok azt mutatják, hogy sokkal dinamikusabbak, mint korábban gondoltak, és permeabilitásukat számos tényező befolyásolhatja, amelyek befolyásolják a hámsejteket.
A gasztrointesztinális epitéliumot számos, funkcionálisan érett, az őssejtek proliferációjából származó sejtek táplálják. Az érett hámsejtek többsége, beleértve a gyomor nyálkahártyáját és a vékonybélben lévő abszorptív sejteket, gyors forgási sebességet mutat, és csak néhány nappal a kialakulásuk után hal meg. Az epitheliális integritás fenntartása tehát pontos egyensúlyt követel a sejtproliferáció és a sejthalál között.
A gasztrointesztinális epithelium folyamatos feltöltését támogató őssejtek a gyomorágyak közepén és a kicsi és vastagburok kriptájában találhatók. A vékonybél epitheliális sejtdinamikáját különösen jól vizsgálták. Ezek az őssejtek folyamatosan szaporodnak, hogy olyan sejteket szolgáltassanak, amelyek ezután abszorptív enterocitákká, mucus-szekretáló pikáns sejtekké, enteroendokrin sejtekké és Paneth sejtekké differenciálódnak. Kivéve a Paneth sejteket, amelyek a kriptákban maradnak, a többi sejtek megkülönböztetik érett formáikat, amikor a kriptákból vándorolnak, hogy kicseréljék a villi csúcsaiból extrudált sejteket. Ez a migráció körülbelül 3-6 napig tart.
Az extrinsic gasztrointesztinális gát
Mucus és Bicarbonate
A teljes gyomor-bélhám-epitélium nyálkahártyával van bevonva, amelyet a hám részét képező sejtek szintetizálják. A bélsár fontos szerepet játszik a hámfeszültség enyhítésében a hámban és számos módon járul hozzá a gátló funkcióhoz. A mucin molekulákban gazdag szénhidrátok kötődnek a baktériumokhoz, amelyek elősegítik az epitheliális kolonizáció megakadályozását, és aggregációt okozva felgyorsítják a clearance-et. A hidrofil molekulák diffúziója lényegesen alacsonyabb a nyálkahártyánál, mint a vizes oldatban, amelyről azt gondolják, hogy késlelteti a különböző károsító anyagok – köztük a gyomorsav – diffúzióját az epithelium felületére.
A nyálkahártyán való bevonás mellett a gyomor és a nyombél hámsejtjei bikarbonát-ionját apikális arcukra bontják. Ez a hólyagos plazmamembrán mentén semleges pH fenntartására szolgál, annak ellenére, hogy a lumenben igen savas körülmények állnak fenn.
Hormonok és citokinek
A gyomor- és bélhámsejtek normális proliferációját, valamint az ilyen károsodásra adott válaszként a proliferációt ismert módon nagyszámú endokrin és parakrin faktor befolyásolja. Ismert, hogy számos bélben oldódó hormon fokozza a proliferáció mértékét. Az epithelium különböző sérülési formái akár fokozott vagy elnyomott sejtproliferációhoz vezethetnek. Például bebizonyosodott, hogy a kutya vékonybélének egy részének reszekcióját hólyagos sejtek hiperpláziája és növekvő villamos hossza követi orálisan beadott állatokban. A parenterálisan táplált állatok nem mutatták ugyanazt a kompenzációs hiperplázist, jelezve, hogy más tényezők mellett a helyi tápanyagok fontos szerepet játszanak a sejtdinamikában.

A prosztaglandinok, különösen a prosztaglandin E2 és a prosztaciklin, régóta ismert, hogy “citoprotektív” hatásai vannak a gasztrointesztinális epitéliumra. Sok emlős esetében gyakori klinikai korreláció az, hogy az aszpirin és más, nem szteroid gyulladáscsökkentő szerek (NSAID-ok) alkalmazása, amelyek gátolják a prosztaglandinszintézist, gyakran társulnak a gyomor eróziójával és fekélyeivel. A kutyák különösen érzékenyek erre a mellékhatásra. A prosztaglandinok az arachidonsavból nyert nyálkahártyán belül ciklooxigenázok hatására szintetizálódnak. A citoprotektív hatásuk úgy tűnik, hogy a nyálkahártya-nyálkahártya és a hidrogénkarbonát szekréciójának stimulálására, a nyálkahártya-véráramlásra és különösen a gyomorra kifejtett hatás komplex képességére vezethető vissza, hogy korlátozza a sav hátsó diffúzióját az epitéliumba. Jelentős erőfeszítések folynak olyan NSAID-ok kifejlesztésére, amelyek nem gátolják a nyálkahártya-prosztaglandinszintézist.
Két olyan peptid, amelyek figyelmet kaptak a barrier-karbantartásban betöltött szerepükre epidermális növekedési faktor (EGF) és transzformáló növekedési faktor-alfa (TGF-alfa). Az EGF nyálban és nyombélben szekretálódik, míg a TGF-alfát gyomor-epiteliális sejtek termelik. Mindkét peptid kötődik egy közös receptorhoz, és stimulálja az epithelialis sejtproliferációt. A gyomorban fokozzák a nyálkaiválasztást és gátolják a savtermelést. Más citokinek, mint például a fibroblaszt növekedési faktor és a hepatocita növekedési faktor kimutatták, hogy fokozzák a gyomor-bélrendszeri fekélyek gyógyulását kísérleti modellekben.
Trefoil fehérjék olyan kis peptidek családja, amelyeket bőségesen választanak ki a gyomor- és bélnyálkahártyán lévő üstsejtek, és bevonják a hámsejtek apikális arcát. Megkülönböztető molekuláris szerkezete úgy tűnik, hogy ellenáll a proteolitikus pusztulásnak. Számos tanulmány kimutatta, hogy a trefoil peptidek fontos szerepet játszanak a nyálkahártya integritásában, a sérülések javításában és az epithelialis sejtproliferáció korlátozásában. Megmutatták, hogy megvédik a hámot a mérgező vegyszerek és gyógyszerek széles skálájától. A trefoil fehérjék központi szerepet töltenek be az epitheliális károsodás javításának visszaállítási fázisában is, ahol a hámsejtek lelapulnak és a sebfelületről áttérnek a denudált területek fedésére. A trefoil géneket célzott delécióval rendelkező egerek túlzottan reagáltak az enyhe kémiai sérülésekre és a késleltetett nyálkahártya-gyógyulásra.
Egy másik olyan molekula, amely meghatározó szerepet játszik a nyálkahártya integritásában és a gátló funkcióban nitrogén-oxid (NEM). Paradox módon a NO is hozzájárul a nyálkahártya-sérüléshez számos emésztőrendszeri betegségben. Ezt a molekulát argininből szintetizáljuk a nitrogén-oxid-szintézis (NOS) három izoformájának egyikével. Az e területen végzett kutatások nagy része az NO donorok, például a glicerltrinitrát vagy a NOS inhibitorok hatásának megértésére összpontosított. Több modellben a NO donorok jelentősen csökkentették a mérgező vegyi anyagok (például az etanol) által okozott nyálkahártya-sérülések súlyosságát, vagy az iszkémia és reperfúzió társultak. Hasonlóképpen, a patkányok gyomorfekélyének gyógyulását NO donorok alkalmazásával gyorsították. Egy másik érdekes megfigyelés az, hogy a NO donorok és az NSAID-k együttes alkalmazása gyulladáscsökkentő tulajdonságokat eredményez, amelyek hasonlóak a nem szteroid gyulladáscsökkentőkhöz, de kisebb mértékben károsítják a gyomor-bélrendszeri nyálkahártyákat. A NOS inhibitorokat olyan helyzetek kezelésére vizsgálják, amelyekben a NO túltermelődik és hozzájárul a nyálkahártya sérüléséhez.
Antibiotikus peptidek és antitestek
A barrier funkció fontos része, hogy megakadályozza a baktériumok átjutását a lumenből az epitéliumon keresztül. A Paneth-sejtek hím epitheliális granulociták, amelyek számos emlős vékonybél-kriptájában találhatók. Számos antimikrobiális peptidet szintetizálnak és titrálnak, köztük az alfa-defenzin izoformáit, amelyek cryptdinekként (“crypt defensin”) is ismertek. Ezek a peptidek antimikrobiális aktivitást mutatnak a potenciális kórokozók számával szemben, beleértve többféle baktériumot, néhány élesztőt és Giardia trophozoitet. A hatásmechanizmusuk valószínűleg hasonló a neutrofil alfa-defenzinhez, amely permeabilizálja a célsejtmembránokat.
A nem specifikus antimikrobiális molekulák mellett a gátló funkciót a gasztrointesztinális immunrendszer támogatja. Ennek a védelmi rendszernek egyik aspektusa a hám nagy részét a szekréciós immunglobulin A. Az antitest ezen osztályát szubepitheliális plazmasejtekből választják ki, és átjutnak az epitheliumon át a lumenbe. A lumenális IgA antigén gátlást biztosít baktériumok és más antigének kötésével. Ez a gátló funkció specifikusan specifikus antigénekre és korábbi válaszfelvételre van szükség a válasz kifejlesztéséhez.
A barrier funkció megzavarása
Annak ellenére, hogy robusztus és sokoldalú természetű, a gasztrointesztinális gátat megszeghetjük. A baktériumok és vírusok által okozott lokális fertőzések, toxinokkal való érintkezés vagy fizikai sérülések, valamint számos szisztémás betegség a zavart okozza. Ilyen problémák lehetnek enyheek és könnyen javíthatók, vagy súlyosak és végzetesek lehetnek.
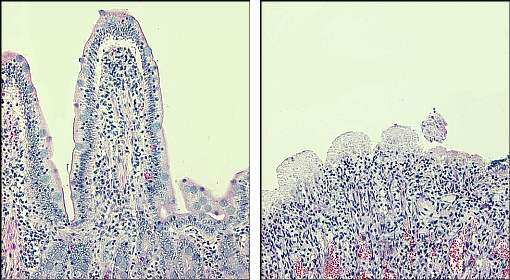
Az alábbi mikrografák a gát súlyos megzavarását mutatják. A bal oldalon a normál kutya vékonybéléből származó nyálkahártya, nagy hólyaggal, melyet az ép hám borít a lumenbe. A jobb oldali kép (ugyanazon nagyítás) a Salmonella bélgyulladásban meghalt kutyából származó vékonybél-nyálkahártyát mutatja – jegyezzük meg a teljesen eldugult epitheliumot és a villi elpusztítását.
Ischaemia és reperfúziós sérülés
Az iszkémia és a reperfúziós sérülés következtében kialakuló gasztrointesztinális gát megrongálódása gyakori és súlyos állapot. Az iszkémia akkor fordul elő, amikor a vér áramlása nem elegendő az oxigén és a tápanyagok mennyiségének a sejt integritásának fenntartásához szükséges mennyiségben. Reperfúziós károsodás következik be, amikor a vér áramlását iszkémiás szövetre állítják vissza.
A gasztrointesztinális iszkémia két alapvető típusú rendellenesség eredménye, melyek mindegyike veszélyeztetheti az epitheliális gátat:
- Nem-occlusi ischaemia olyan szisztémás állapotok, mint a keringési sokk, szepszis vagy szívelégtelenség.
- Nyugtató iszkémia olyan állapotokra utal, amelyek közvetlenül gátolják a gyomor-bélrendszeri véráramlást, például fulladást, volvulust vagy thromboemboliát.
A gyomor-bél rendszeri reperfúziós sérülése, különösen a nyálkahártya, elsősorban a reaktív oxigénfajok, köztük a szuperoxid, a hidrogén-peroxid és a hidroxilgyökök képződésének tulajdonítható. Ezek az oxidáló szerek a nyálkahártyán belül, valamint az ischaemia folyamán aktivált számos helyi leukocitában keletkeznek.
A reperfúzió során keletkező oxigénből származó szabadgyökök olyan eseménysorozatot indítanak el, amely nyálkahártya-sérülést és a gát megzavarását okozza. Közvetlenül károsítják a sejtmembránokat lipid peroxidok kialakításával, ami számos foszfolipidből (például vérlemezke-aktiváló faktorból és leukotriénekből) származó gyulladásos mediátorok előállításához vezet. Ezek a gyulladásgátló szerek kemoattraktánsok a neutrofilek számára, amelyek a nyálkahártyába vándorolnak, felszabadítják saját reaktív oxigénmetabolitjaikat, és további károkat okoznak az intrinsic epithelialis gáton. Az iszkémia eredetileg kicsi hatását ezáltal erősíti a barrier funkció jelentős mértékű károsodása. Továbbá a gyomor-bél traktusban előidézett gyulladásos mediátorok károsíthatják a távoli szöveteket, ami szisztémás betegséghez vezethet.
Az iszkémia-reperfúziós károsodás megfigyelt hatása a megnövekedett vascularis permeabilitás és ennek következtében a subepitheliális ödéma, a hámsejtek és a villamosok tömeges elvesztése. Még a viszonylag enyhe károsodás is megakadályozza a gátló funkciót, és a baktériumok és toxinok transzlokációjához vezethet a lumenből a szisztémás keringésbe. A kaszkád károsodásának megakadályozása érdekében számos kezelést fejlesztettek és teszteltek, ideértve az antioxidánsok, például a szuperoxid-diszmutáz alkalmazását és a vérlemezke-aktiváló faktor antagonisták alkalmazását a gyulladásos mediátorok hatásának blokkolására.
Neutrofilek és mucosális sérülések
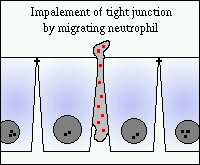
A bélrendszeri nyálkahártyák sokfélesége, beleértve a fertőző folyamatokat, az iszkémia és a káros vegyületeket, elősegítik a neutrofilek beszivárgását. Ez a közös végpont eredménye, mivel sokféle sérülés vezet a neutrofil kemoattraktánsok helyi előállításához, mint például a leukotriének, az interleukinok és az aktivált komplement komponensek. A kemoattraktánsokra reagálva a neutrofilek átjutnak a kapillárisokból, beszivárognak a subepithelialis nyálkahártyában, és gyakran átjutnak a gyomor- vagy bélhámon keresztül. Az epitélium keresztezésénél a neutrofileknek meg kell szakítani a hámsejtek közötti junkcionális komplexeket. Ez a “behatás” a szoros csomópontokon keresztül szükségszerűen átmeneti megnövekedést eredményez az elfogadhatóságban. Ha a sérelem kisebb, akkor a csomópontok gyorsan visszazáródnak, de a nagyszámú neutrofil sejtek átvivése jelentős mértékben károsítja a gátló funkciót.
A stressz hatásai
A stressz számtalan formában jön létre, és szerves része az összes betegségnek és traumának. A stresszválasz szó szerint több tucat hormon és citokin modulációját, valamint a neurotranszmisszióra gyakorolt jelentős hatásokat foglalja magában. Azonban, a stressz legfontosabb hatása a gyomor-bél traktusra, hogy csökkenti a nyálkahártya véráramlását, és ezáltal veszélyezteti a nyálkahártya-barrier integritását. Többek között a csökkent nyálkahártya-véráramlás megszünteti a nyálkahártyák termelését, és korlátozza a protonok diffúzióját. Ennek következtében a jelentős stressz szinte mindig a nyálkahártya eróziójához kapcsolódik, különösen a gyomorban. Ezen elváltozások többsége szubklinikus, de a gyomor-bélrendszeri vérzés és a szepszis nem ritka következmények.
A cukorbetegség és az elhízás jól ismert kockázati tényezők a GI gát és a hosszan tartó hyperglykaemia ezekhez a rendellenességekhez társuló elsődleges mechanizmus lehet az ilyen zavarok elsődleges mechanizmusa.
Visszaállítás és gyógyulás sérülés után
A gyomor-bélhám epiteliájának megzavarását követő kritikus első feladat az, hogy lefedje a denaturált területet, és újból létrehozza a belső gátat. Az epitél gyors helyreállítását olyan eljárással végezzük, melyet visszaállítunk – a hólyagos sejtek a hiba szomszédságában szétterülnek és a kioltott aljzatmembrán fölött vándorolnak. A vékonybélben ezt a folyamatot az érintett villi gyors összehúzódása és lerövidítése segíti elő, ami csökkenti a pincében lévő membrán területét, amelyet le kell fedni.
A restitution gyors mechanizmust biztosít a hiba hiányának fedezésére, és nem jár az epithelialis sejtek proliferációjával. Ez egy olyan területet eredményez, amely védett, de nem élettani szempontból funkcionális. A gyógyítás megköveteli, hogy a hiba peremén lévő hámsejtek szaporodjanak, differenciálódjanak és migrálódjanak a sérült területre, hogy visszaállítsák a normál sejtes felépítést és funkciót.
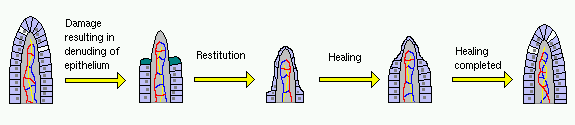
Úgy tűnik, hogy a restitúció számos többnyire parakrin szabályozó által stimulált. A folyamatban a helyi prosztaglandinok és az alvadréteg fehérjék nyilvánvalóan érintettek, és termelésük elnyomása jelentősen késlelteti a helyreállítást. A restitúcióban részt vevő molekulák egy másik csoportja a poliaminok, például a spermium, a spermidin és a putreszin. Ezek a molekulák számos étrendben megtalálhatók, és a gasztrointesztinális nyálkahártyák szintetizálják őket. A poliaminok enterális adagolását kísérletes modelleken mutatták be, hogy felgyorsítsák a nyálkahártya elváltozások visszaállítását és gyógyulását.
Referenciák és vélemények
- Allaire JM, Crowley SM, Law HT, stb. A bélhám: a nyálkahártya-immunitás központi koordinátora. Trends Immunol 2018, DOI: https://doi.org/10.1016/j.it.2018.04.002
- Blikslager AT, Roberts MC: A bélnyálkahártya javításának mechanizmusa. J Am Vet Med Assoc 211: 1437-1441 (1997).
- Dieckgraefe BK, Stenson WF, Alpers DH: Gasztrointesztinális epitheliális válasz a sérülésre. Curr Opin Gastroenterol 12: 109-114, 1996.
- Engle E, Guth PH, Nishizaki Y, Kaunitz JD: A gyomornyálkahártya gátló funkciója. Am J Physiol 269: G994-999, 1995.
- Filep J, Herman F, Braquet P, Mozes T: A vérben lévő vérlemezke-aktiváló faktor fokozott szintje a kutya belében lévő iszkémia után. Biochem Biophys Res Commun 158, 353-355 (1989)].
- Gayle JM, Blikslager AT, Jones SL: A neutrofilek szerepe az intesztinális nyálkahártya-sérülésben. J Am Vet Med Assoc 217: 498-500, 2000.
- Kubes P, Hunter J, Granger ND: Ischaemia / reperfúzió által kiváltott macska bélműködés: a granulocita felvétel fontossága. Gastroenterology 103, 807-812 (1992).
- Lichtenberger LM: A gasztrointesztinális nyálkahártya hidrofób gátló tulajdonságai. Ann Review Physio 57, 565-583 (1995)].
- Mashimo H, Wu D, Podolsky DK, Fishman M: A bélnyálkahártya gyengébb védelme egerekben, amelyekben nincs bélflós faktor. Science 274, 262-264 (1996)].
- Murphy MS: növekedési faktorok és a gyomor-bél traktus. Nutrition 14, 771-774 (1998).
- Muscara MN, Wallace JL: Nitrogén-oxid V. Nitrogén-oxid donorok és inhibitorok terápiás potenciálja. Am J Physiol 276: G1313-1316, 1999.
- Thaiss CA, Levy M, Grosheva I, stb. A hyperglykaemia a bélgátló zavar és a bélfertőzés veszélyének a következménye. Science 359: 1376-82, 2018.
- Thompson JS: A kritikus betegségre adott bélreakció. Am J Gastroenterology 90: 190-200, 1995.
- Wallace JL, Miller MJ: Nyálkahártya-védelemben levő nitrogén-oxid: egy kicsit hosszú utat mutat. Gastroenterology 119, 515-520 (2000).
Gyomor: Bevezetés és index ![]()
Frissítve 2018. május. Megjegyzések küldése Richard.Bowen@colostate.edu címre
Az oldal hindi fordítását Nikol Barton hozta létre, és elérhető a hindi fordításban
Vér-prosztata-gát

A Journal of Urology

Bevezetés:
Vér-epitéliális korlátokat írtak le a herékben és az epididimusban, de az ilyen akadályok lehetősége a férfi ártéri traktus más területein nem kapott alapos vizsgálatot. Ennek a vizsgálatnak az a célja, hogy in vivo mikro-punktúrát használjon annak megállapítására, hogy a vér-epiteliális gát fennáll-e a patkány ventrális prosztata formájában. Ezen túlmenően, prosztata gyulladásos modell alkalmazásával arra törekedtünk, hogy megvizsgáljuk a gyulladás hatását a vérben hordozó molekulák és leukociták átjutására a prosztata ductalis lumenbe.
Anyagok és metódusok:
A felnőtt Sprague-Dawley patkányokat két csoportra osztottuk, kontroll és 24 órás lipopoliszacharid (LPS) indukálta gyulladás. Mindkét csoportot radioaktívan jelölt 3 H-dextrán, 14 C karbamid és 3 H vizet tartalmazó vaszkuláris infúziónak vetettük alá. A prosztata ductalis folyadék (DF) és az artériás vér egyidejű in vivo mikropunktúrás mintavétele többszörös 120 percen keresztül történt. A radioaktívan jelzett vegyületek transzsztitéli mozgását minden egyes mintavételi időpontban kvantáltuk a DF izotóp-koncentrációk expressziójával a szérum izotópkoncentráció százalékában az adott időpontban. A kontroll és a gyulladt prosztata reprezentatív reprezentatív mintáinak szövettana a gyulladásos válasz megerősítésére és a leukociták ductalis lumenbe való bejutásának vizsgálatára szolgál.
A radioaktívan jelölt vegyületek transepiteliális mozgása a vérből a prosztata lumenbe közvetlen kapcsolatban áll a vegyület molekulatömegével. 3 H-víz (MW = 18) mozgása a ductalis lámpánál viszonylag gyors volt a szérum értékek 70-80% -ában. 14 C karbamidot (MW = 60) a ductalis folyadék (50-60% szérum értékek) és 3 H dextrán (MW = 2 × 10 6) közbülső penetrációja lényegében kizárt a beléptetésből (
Előző cikk
Következő cikk

Kommentek
Kommenteléshez kérlek, jelentkezz be: